Разделы сайта
Иннервация клетки-мишени
Механизмы управления конусов роста, описанные до сих пор, направляют аксон к его конечной цели. Однако проблема того, как каждый аксон подходит к своей клетке-мишени, остается открытой. Путь, по которому ганглиозные клетки сетчатки иннервируют свои клетки-мишени в области передних бугров покрышки, является примером того, какие сигналы обеспечивают достижение правильного паттерна иннервации. Во время развития аксоны ганглиозных клеток задней или височной (temporal) части сетчатки направляются для иннервации в переднюю часть тектума, а клетки передней (nasal) части сетчатки направляются для иннервации задней части тектума (рис. 2).
|
|
|
Рис. 1CD. (С и D) Семафорим III (Sema3A) — хемореппелент дальней дистанции для сенсорных аксонов. Спинальный ганглий эмбриона крысы (слева на каждой панели) культивировали в течение 48 часов рядом с агрегатами COS клеток. Белыми кружками показаны контуры ганглия. Для того чтобы вызывать рост сенсорных волокон малого диаметра в среду был добавлен ФРН. (С) Контрольные COS клетки. Аксоны растут в виде венчика. (D) COS клетки, секретирующие рекомбинантный Sema3A. Sema3A отталкивает аксоны. |

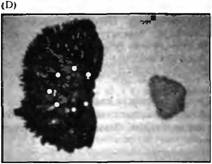
В элегантной серии экспериментов Бонхоеффер с коллегами продемонстрировали, что аксоны определяют территории своей иннервации посредством отталкивающих взаимодействий, которые предотвращают вторжение аксонов височной зоны в область задней части тектума. Ганглиозные клетки височной сетчатки были помещены в культуру, где рядом с ними находились поверхности, покрытые мембранами, выделенными либо из передней, либо из задней области тектума. В этих условиях аксоны отталкивались мембранами из задней части тектума и росли преимущественно в направлении мембран своих естественных мишеней, мембран передней части тектума. Любопытно, что аксоны сетчатки, не имея подобного выбора, быстро растут в направлении любого субстрата, как из передней, так и из задней части тектума.
Молекулы, ответственные за такие отталкивающие взаимодействия, принадлежат к семейству рецепторов, связанных с тирозинкиназой (известных как Eph киназы), а также их лигандов (называемых эфринами, ephrines). Эфрин-А2 и эфрин-5 экспрессируются в тектуме во время образования ретинотектальных связей, и их концентрация постепенно увеличивается в направлении спереди назад. Рецептор к Eph-АЗ экспрессируется на аксонах клеток сетчатки соответственно назотемпоралыюму градиенту. При заключении в липидные пузырьки и добавлении в среду, омывающую аксоны клеток височной части сетчатки, эфрины А2 и А5 вызывают отделение конусов роста от субстратов и их ретракцию. Эфрины и семейство Eph рецепторов, связанных с тирозинкиназой, действуют во всех частях развивающейся нервной системы, влияя на нахождение пути аксоном, миграцию клеток, образование связей между ними, в основном демонстрируя сходный отталкивающий механизм.
Одного рострокаудального градиента недостаточно для того, чтобы аксоны клеток сетчатки правильно достигли своего окончательного месторасположения в тектуме. Более поздние изменения в рострокаудальных проекциях основываются как на определенно расположенных химических сигналах, так и на механизмах, зависящих от активности нейронов
nextbiology.ru All Rights Reserved. 

