Разделы сайта
Механизм генерации и распространения потенциала действия
Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны.
Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток, называется потенциалом действия.
Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток.
На основе обобщения большого экспериментального материала было установлено, что потенциалы действия возникают в результате избыточного по сравнению с покоем диффузии ионов натрия из окружающей жидкости внутрь клетки.
Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обусловливает восстановление исходного потенциала покоя.
Потоки приблизительно равны по величине, но сдвинуты во времени. Благодаря этому сдвигу во времени и возможно появление потенциала действия.
Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности.
Распространение потенциала действия обусловлено возникновением локальных токов, циркулирующих между возбужденным и невозбужденным участками клетки.
В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя - отрицательный.
В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рисунок).
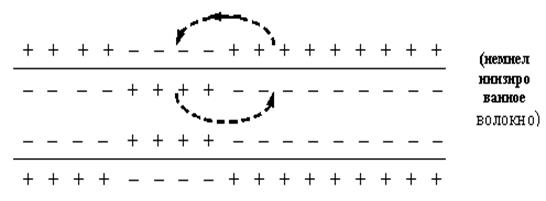
В результате этого между возбужденным и невозбужденным участками мембраны (B и H) имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении.
Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя.
Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время уже происходят восстановительные процессы реполяризации. Вновь возбужденный участок, в свою очередь, становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок.
Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях. В нервной системе прохождение импульсов только в определенном направлении обусловлено наличием синапсов, обладающих односторонней проводимостью.
Под влиянием локальных токов волна возбуждения распространяется вдоль волокна без затухания (бездекрементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения.
Скорость уменьшения мембранного потенциала до критического уровня зависит от разности потенциалов между возбужденным и невозбужденным участками и от кабельных свойств волокна: электрической емкости и сопротивления мембраны, сопротивлений аксоплазмы и окружающей среды.
nextbiology.ru All Rights Reserved. 

